IL-33とアレルギー
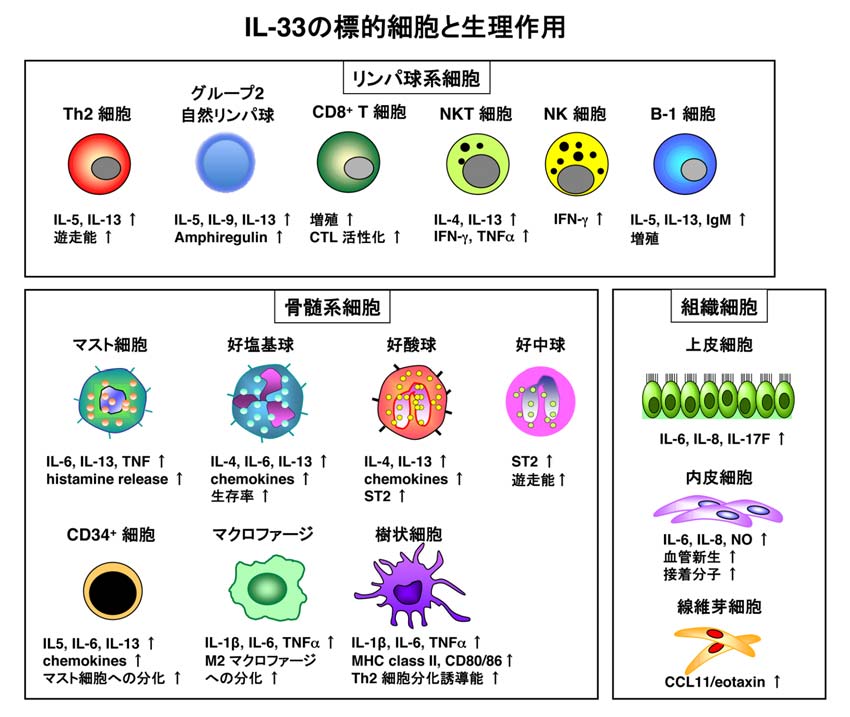
IL-33はIL-1βやIL-18と相同性の高いアミノ酸配列を有するIL-1 ファミリーに属するサイトカインとして、2005年にクローニングされました。その受容体は1989年に発見され、長いあいだそのリガンドが不明だったST2です。IL-33は様々な臓器の上皮細胞や血管内皮細胞の核内に局在し、組織傷害によって短時間で細胞外に放出されます。しかし、その産生機序は未だ不明な点が多いのが現状です。IL-33は好塩基球やマスト細胞以外に、近年相次いで報告されたグループ2自然リンパ球(Group 2 innate lymphoid cells: ILC2s)を刺激してTh2サイトカインを産生することで、「自然型アレルギー」に関与します。
一方、IL-33は抗原と共にTh2細胞を刺激してIL-5/IL-13産生を増強し、「獲得型アレルギー」の発症・増強にも関与します。すなわち、IL-33は「自然型アレルギー」と「獲得型アレルギー」に極めて重要な調節因子であり、それぞれのアレルギー疾患に対する治療標的として期待されています(IL-33の総説 Cytokine Frontiers, 2013)。
IL-33による自然型アレルギーの誘導
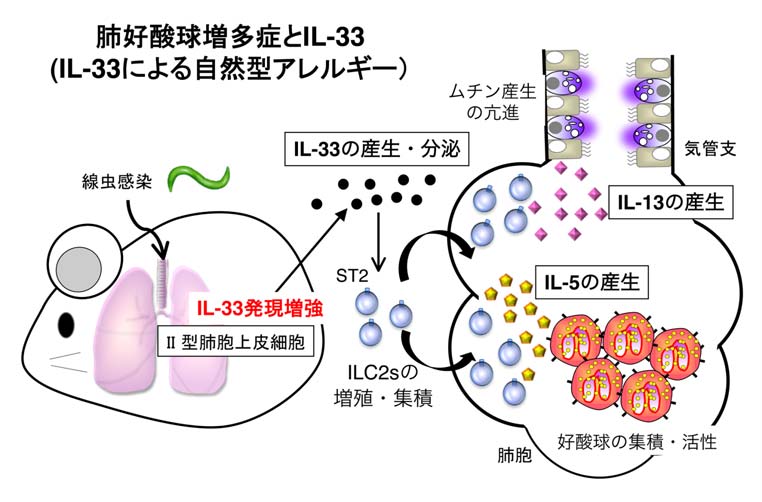
肺好酸球増多症は、末梢血中の好酸球の増加と肺の異常陰影を示し、激しい喘息発作を発症します。寄生虫やカビが原因と考えられていますが、未だ不明な点が多い疾患です。私たちは、正常マウスに腸管寄生線虫を感染することによって肺に著明な好酸球増多を誘導する肺好酸球増多症(Loeffler症候群)のモデルマウスを作製しました。このモデルマウスを用いて、肺好酸球増多症の発症機序を解析し、以下のことを明らかにしました(PNAS, 2013)。
正常マウスは腸管寄生線虫(ヴェネズエラ糞線虫)に感染すると、
- II型肺胞上皮細胞から迅速(1日以内)にIL-33蛋白が産生される。
- このIL-33の作用によって、グループ2自然リンパ球が肺に著明に集積・増加する。
- IL-33はIL-33受容体(ST2)を発現するグループ2自然リンパ球に作用し、多量のIL-5とIL-13を産生する。その結果、
- IL-5は好酸球を肺に集積・活性化を、IL-13は粘液ムチン産生と好酸球遊走を亢進し、感染後3~4日目には好酸球性肺炎を発症する。更に、
- T細胞とB細胞を欠損するRAG2欠損マウスに線虫を感染しても好酸球性肺炎を発症しするのに対し、
- IL-33欠損マウスでは好酸球浸潤は著明に減少した。
尚、インフルエンザ感染に伴う喘息の発症にも、IL-33—グループ2自然リンパ球を介した自然型喘息が関与することも明らかにされています。
IL-33による獲得型アレルギーの発症・増強
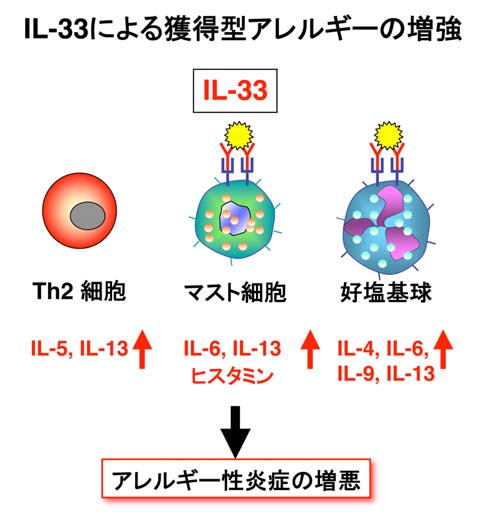
IL-33は抗原と共にTh2細胞を刺激してIL-5/IL-13産生を増強します。また、IL-33はアレルゲンとIgE抗体で活性化された好塩基球とマスト細胞に作用するとTh2サイトカインとヒスタミン産生を増強します。この結果、IL-33は獲得型アレルギー炎症を発症・増強します。
私たちは、ブタクサ花粉特異的アレルギー性鼻炎モデルマウスと経皮感作食物アレルギーモデルマウスを作成し、それぞれのアレルギー症状(鼻炎・アナフィラキシーショック)はIL-33依存的に発症することを明らかにしました。
